Migration of Birds
Origin and Evolution of Migration
The origin and evolution of bird migration has been discussed in ornithological
literature for centuries. As we have seen from the foregoing discussion,
migration exists in many forms throughout the world and probably arose
to satisfy many different needs in different orders of birds at the same
time. New patters, traditions, and routes are arising today as well as
disappearing. Currently, the migration patterns we see are a composite
result of historic influences mixed with present day influences. Even though
the migration of several different species may be very similar, the patterns
exhibited today can be the result of quite different evolutionary processes.
Because it cannot be substantiated by experimental facts, any explanation
of how a particular pattern or route originates is pure conjecture.
The general anatomical and physiological attributes of birds enable
them to develop more diverse and spectacular migratory behavior than any
other group of animals. Their potential for long sustained flights is of
primary importance in pre-adapting birds to successful migrations. Migration
has long since become a definite hereditary habit of many species of birds
that recurs in annual cycles, evidently because of physiological changes
which prompt a search for an environment suitable for reproduction and
survival. Like the bird's other habits its migratory behavior is just as
characteristic as the color of its plumage and, like it, evolved through
natural selection because it was advantageous for the survival of the population.
Its origin has been thought by some to be a mystery locked in past ages,
but by study of the history of how birds came to occupy their present ranges,
information becomes available which suggests theories that may be developed
and explored. Two that are commonly mentioned are termed the "Northern
Ancestral Home Theory" and the "Southern Ancestral Home Theory."
According to the former of these hypotheses, in earlier ages when
conditions of climate, food, and habitat were favorable for existence of
birds throughout the year much further north than is the case today, many
species remained in these nothern latitudes as permanent residents. Today,
such conditions are found only in more southern regions where migrations
are much shorter or nonexistent. Gradually, however, in the Northern Hemisphere
the glacial ice fields advanced southward, causing a southward movement
of conditions favorable to northern birds, until finally all bird life
was confined to southern latitudes. As the ages passed, the ice cap gradually
retreated, and each spring the birds whose ancestral home had been in the
North moved in again to fill newly opened breeding habitat only to be driven
south again at the approach of winter. As the size of the ice-covered area
diminished, the journeys to the summer breeding areas became even longer
until eventually the climatic conditions of the present age became established,
and with them, present patterns of the annual advance and retreat we call
migration.
The opposing theory is simpler in some respects and supposes the
ancenstral home of Northern Hemisphere migratory birds was in the Tropics.
As all bird life tends to overpopulation, there was a constant effort of
young individuals to pioneer and seek breeding grounds where competition
was less severe. Species better adapted to more northern latitudes moved
in that direction for nesting but were kept in check by the glacial ice
and forced to return southward with the recurrence of winter conditions.
Gradually, as the ice retreated, vast areas of virgin country became successively
suitable for summer occupancy, but the winter habitat in the South remained
the home to which the birds returned after the nesting season.
The above two theories presume that the Quaternary glaciations,
which occurred 10,000 to 1 million years ago, have been the predominate
influence on bird migration in North America and Europe as we observe it
today. There is no doubt these extreme climatic and ecologic barriers played
a part in shaping or modifying some patterns, but as Moreau (1951) has
pointed out, well-developed migrations occur in parts of the world, including
the Southern Hemisphere or even within the tropics, where continental glaciation
was not a factor. Furthermore, migrations to fit various needs have probably
been going on ever since birds could fly. The tremendous Pleistocene glaciations
actually occupied less than a hundreth of the time birds have existed on
the earth and probably only determined the details of migrations as we
see them today (Moreau 1951).
The northern and southern ancestral home theories appear diametrically
opposed to each other but Dorst (1963) concludes they are perfectly compatible.
Since the phenomena probably occurred simultaneously, northern migrants
then originated from two stocks: the North Temperate Zone birds sought
refuge to the south during the glacial periods and the tropical avifauna
expanded their range during the interglacial periods. Dorst also stated
this double origin is more prevalent in North America where the tropical
element is most abundant. Birds representing this element include hummingbirds,
tyrant flycatchers, orioles, tanagers, and blackbirds. At some latitudes,
they nest in the same area as the shorebirds which are of arctic parental
stock.
These theories assume migration is a genetic, inherited character,
but we now know in some species it can be modified in the lifetime of one
individual and the place some individuals return to nest or winter is not
the ancestral home but a place to which they had been transported at an
early stage in their development. Traditions that have lived for countless
generations may die overnight if experienced individuals are lost or no
longer active (Hochbaum 1955); migration patterns remain constant only
as long as the factors influencing these patterns remain constant. But
the landscape and the interacting ecological stresses are forever changing,
and we would expect the adaptive behavior of birds to respond with them.
One of these responses to an expanding habitat is colonization of new territory
and expansion of a species' range with accompanying development of a migratory
habit. The search for favorable conditions under which to breed in summer
and to feed in winter, as influenced by competition for space, has been
the principal factor underlying the extension of ranges, usually by young,
nonconditioned individuals. This is exemplified by the northward extension
in historic times of a number of species. Many of these range extensions
have closely followed man's settlement of the area and the subsequent changes
in habitat that man has made.
From the previous descriptions of migration patterns and routes,
it will be observed that the general trend of migration in most northern
populations of North American birds is northwest and southeast. Eastern
species tend to extend their ranges by pushing westward, particularly in
the North. For example, in the Stikine River Valley of northern British
Columbia and southwestern Alaska the common nighthawk, chipping sparrow,
rusty blackbird, yellow warbler, American redstart, and others have established
breeding stations at points 20 to 100 miles from the Pacific Ocean. The
northern race of the American robin, common flickers, dark-eyed juncos,
blackpoll warblers, yellow-rumped warblers, and ovenbirds, all common eastern
species, also are established as breeding birds in western Alaska. The
ovenbird has even been detected on the lower Yukon River, and the sandhill
crane and gray-checked thrush have moved across Bering Strait into Siberia.
These birds continue to migrate through the eastern part of the continent.
Instead of taking the shortest route south, they retrace the direction
of their westward expansion and move southward along the same avenues as
their more eastern relatives.
The red-eyed vireo is essentially an inhabitant of states east
of the Great Plains, but an arm of its breeding range extends northwest
to the Pacific coast in British Columbia (Fig. 30). It seems evident this
is a range extension that has taken place comparatively recently by a westward
movement via deciduous woodland corridors, and the invaders retrace in
spring and fall the general route by which they originally entered the
country.
In the case of the bobolink, a new extension of the breeding range
and a subsequent change in the migration of the species has taken place
since settlement by European man (Fig. 19). Because the bobolink is a bird
of damp meadows, it was originally cut off from the Western States by the
intervening arid regions, but with the advent of irrigation and the bringing
of large areas under cultivation, small colonies of nesting bobolinks appeared
at various western points. Now the species is established as a regular
breeder in the great mountain parks and irrigated valleys of Colorado and
elsewhere almost to the Pacific coast. These western pioneers must fly
long distances east and west to reach the western edge of the route followed
by the bulk of the bobolinks that breed in the northern United States and
southern Canada.
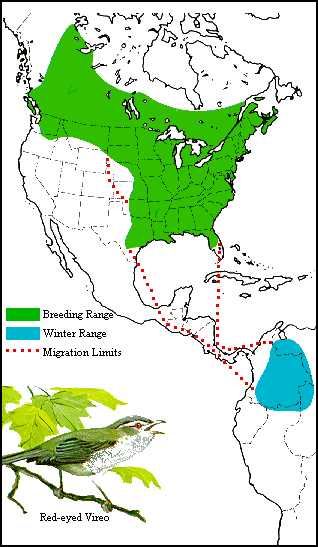 |
| Figure 30. Distribution and migration of the red-eyed vireo. It is
evident that the redeyed vireo has only recently invaded Washington by
an extension of its breeding range( almost due west from the upper Missouri
Valley. Like the bobolink (Fig. 19), however, the western breeders do not
take the short cut south or southeast from their nesting grounds but migrate
spring and fall along the route traveled in making the extension. |
During the past few decades, various populations of Canada geese have
altered their migration patterns as a result of transplanting brood stock,
development of refuges or changing agricultural practices. These routes
will continue to change in the coming years as long as these factors are
in a state of flux. It has been shown that man can establish breeding colonies
of Canada geese with young birds almost anywhere.
Europe also has several good examples of changes in migration
routes through range extension. One of the best examples is the serin.
During the past century, this European finch has spread its breeding range
from around the Mediterranean Sea to include the entire continent. While
the Mediterranean populations remain sedentary the more northern breeding
birds are migratory. Most likely, those birds that did not migrate from
the North were eliminated by severe weather. Similarly, the wheatear, yellow
wagtail, and Arctic warbler have extended their breeding ranges eastward
across the Bering Sea into Alaska, but the wheatear, for instance, migrates
all the way back across Asia to Africa where it winters with other wheatears
coming from Europe, Iceland, and Greenland.
As bird populations become more and more migratory, we might expect
their flight capabilities to be enhanced accordingly. These changes in
morphology are readily seen in wing shape. Several groups of birds have
closely related species or populations some of which are migratory and
others sedentary. The sedentary species or populations have more rounded
wings because of the relative length of the wing quills. On the other hand,
populations that migrate great distances, such as albatrosses, falcons,
swifts, various shorebirds and terns, have more pointed wings. Kipp ( 1942,
1958) demonstrated this using orioles. The sedentary black-headed oriole
of India has a well-rounded wing whereas the closely related black-naped
oriole is migratory between India and Siberia and has primaries that are
much more pointed and well developed.
Thus it seems the origin and evolution of migration have roots
in the present that are deep in the past. The important thing to consider
in the evolution of a migratory trait is whether a population can adapt
to new conditions by genetic modification of its physiology and habits.
The migratory habit has evolved in those populations in which, on the average,
more individuals survive by moving to a different area part of the year
than if they remained in the same area all year.
Previous Section -- Patterns of Migration
Return to Contents
Next Section -- Where We Stand